

- 📖 Geeky Medics OSCE Book
- ⚡ Geeky Medics Bundles
- ✨ 1300+ OSCE Stations
- ✅ OSCE Checklist PDF Booklet
- 🧠 UKMLA AKT Question Bank
- 💊 PSA Question Bank
- 💉 Clinical Skills App
- 🗂️ Flashcard Collections | OSCE, Medicine, Surgery, Anatomy
- 💬 SCA Cases for MRCGP
To be the first to know about our latest videos subscribe to our YouTube channel 🙌
Introduction
The central nervous system uses ascending and descending pathways to communicate with the external environment, this article will discuss the descending (motor) tracts.
Descending tracts carry motor information in efferent nerves from upper motor neurons of cortical structures like the cerebellum and cerebrum. The descending tracts transmit this information to lower motor neurons, allowing it to reach muscles.
Efferent is derived from the Latin ‘effero’ which means ‘to carry out.’
With each of the pathways we discuss, there are two neurons present:
- Upper motor neurons (UMN): brain and brainstem to the ventral horn of the spinal cord
- Lower motor neurons (LMN): from the ventral horn of the spinal cord to the peripheral muscles
We typically describe the neurons in the descending tracts as being UMNs, and those leaving the spinal cord as LMNs. This will become particularly relevant during the discussion of pathologies that present in the motor pathways.
There are many motor tracts in the spinal cord. Some of these are under conscious control and others under unconscious, reflexive or responsive control. These motor tracts can be grouped functionally into pyramidal and extrapyramidal tracts.
These functional groups contain several anatomical tracts, one for each side of the body:
- Pyramidal: conscious control of muscles from the cerebral cortex to the muscles of the body and face; and
- Extrapyramidal: unconscious, reflexive or responsive control of muscles from various brainstem structures to postural or anti-gravity muscles
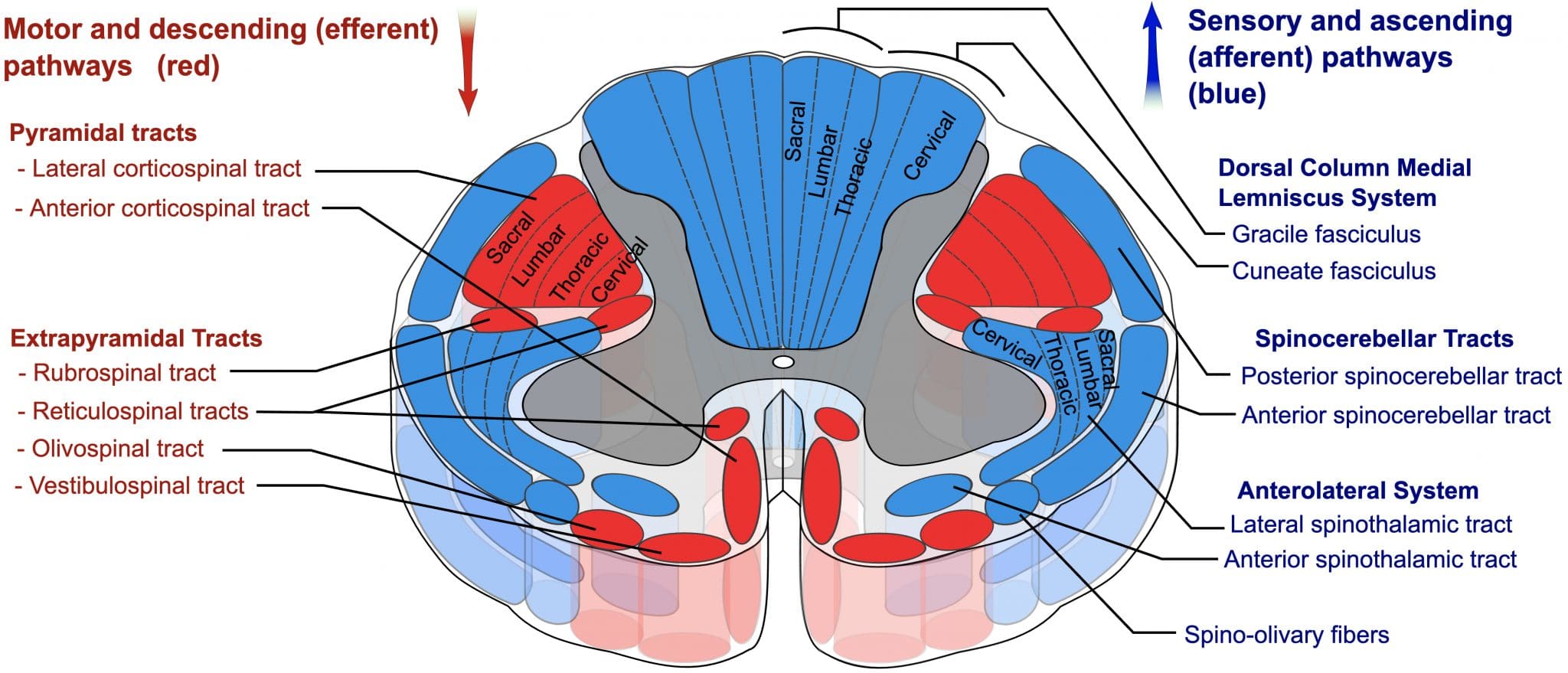
Pyramidal tracts
The pyramidal tracts are named as such due to their course through the pyramids of the medulla oblongata. The pyramidal tracts are responsible for the conscious, voluntary control of the body and face muscles.
They can be divided into two tracts that supply each of these areas:
- Corticospinal tract: cortex to spine (body)
- Corticobulbar tract: cortex to ‘bulb’ (Latin for ‘swelling or bulb of an onion’ like the head and neck)
Corticospinal tract (CST)
The CST communicates with three major cortical areas:
- Primary motor cortex: located in the precentral gyrus; execution of movements
- Premotor cortex: responsible for the control of behaviour, particularly of the trunk muscles
- Supplementary motor cortex: stabilisation and coordination of the body during bimanual movements
The fourth cortical area the CST communicates with is the posterior parietal cortex for integration with and modulation of incoming sensory information.
Neurons exiting the cerebral cortex in one of the three major regions above converge to form the white matter structure in the brain known as the internal capsule. The internal capsule is located between the basal ganglia and thalamus; two highly vascularised structures in the deep brain.
After passing through the internal capsule, the fibres continue to pass down through the centre of the crus cerebri of the midbrain, before entering the pons and medulla.
As the CST passes through the caudal medulla, it divides into the lateral and anterior corticospinal tracts:
- Lateral CST: decussate in the pyramid of the medulla
- Anterior CST: stay ipsilateral
These tracts then descend into the spinal cord, terminating in the ventral horn of the spinal cord where they synapse onto LMNs to supply the peripheral musculature.
The anterior CST remains ipsilateral and descends only to the cervical and upper thoracic spinal cord, where they decussate at the level of the nerve root they supply.
Table 1. The corticospinal tract.
| UMN/LMN | Structure | Ipsilateral | Contralateral |
| UMN | Cerebral cortex | X | |
| UMN | Internal capsule | X | |
| UMN | Crus cerebri | X | |
| UMN | Pons | X | |
| UMN | Rostral medulla | X | |
| UMN | Caudal medulla (pyramids) | aCST | LCST |
| UMN | Spinal cord | aCST | LCST |
| LMN | Level of nerve root | aCST + LCST | |
| LMN | Peripheral nerve | X |

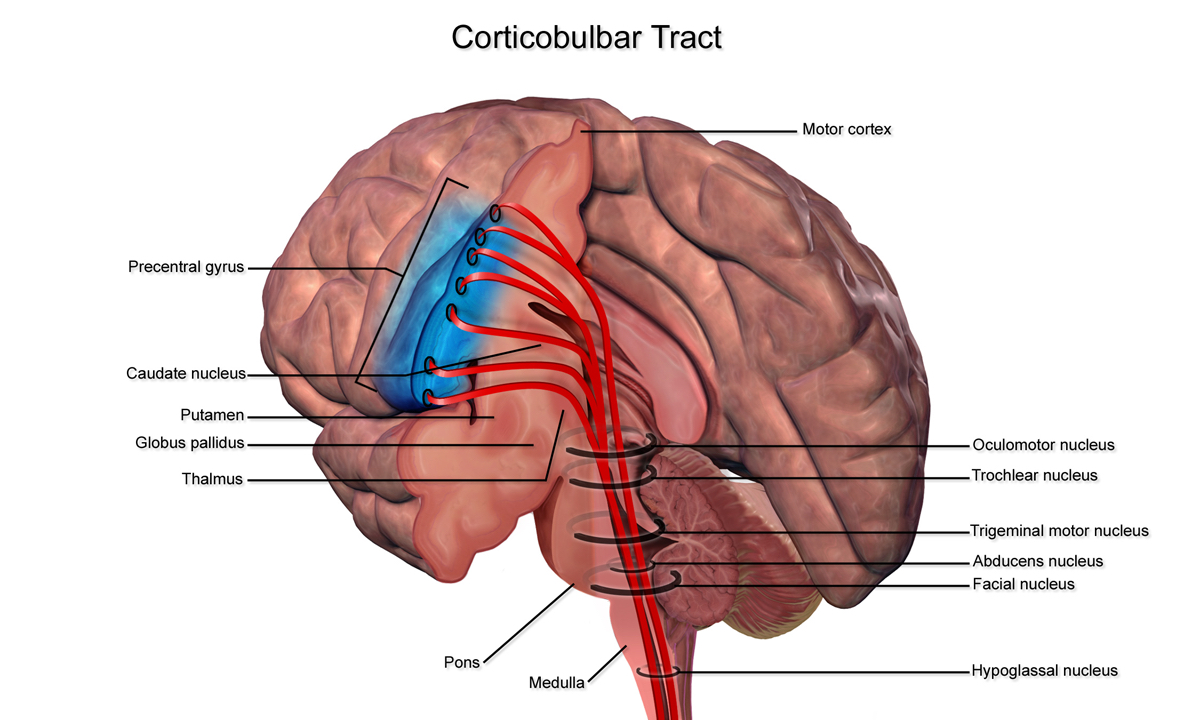
Corticobulbar tracts (CBT)
Arising from the lateral aspect of the primary motor cortex (the cephalic region of the motor homunculus), the CBTs receive mostly the same inputs as the CSTs.
They follow a similar path but terminate in the brainstem at the motor nuclei rather than continuing down to the spinal cord.
In the brainstem, they synapse on the cranial nerve motor nuclei, which are LMN structures that supply the head and neck muscles.
It is important to understand the clinical implication of damage to the CBTs. The UMN structures of the CBT provide bilateral innervation to the structures of the head and neck. This is the case for all head and neck muscle cranial nerve nuclei except:
- Cranial nerve VII, which derives only contralateral UMNs for the lower half of the face
- Cranial nerve XII, which derives only contralateral UMNs for the tongue
Table 2. Cranial nerve nuclei.
| Cranial nerve nucleus | Ipsilateral UMN | Contralateral UMN | |
| V | X | X | |
| VII | Upper half | X | X |
| Lower half | – | X | |
| XI | X | X | |
| XII | X | X | |
Extrapyramidal tracts
The extrapyramidal tracts all originate in the brainstem and do not pass through the pyramids.
These tracts all carry motor fibres to the spinal cord that allow for unconscious, reflexive or responsive movement of muscles to control balance, locomotion, posture and tone.
There are four tracts:
- Reticulospinal
- Vestibulospinal
- Rubrospinal
- Tectospinal
Reticulospinal tracts
The reticulospinal tracts do not decussate.
There are two reticulospinal tracts:
- Medial reticulospinal tract: originates in the pons and contributes to voluntary movements and increases in muscle tone in response to alerting or activating stimuli that stimulate the reticular activating system;
- Lateral reticulospinal tract: originates in the medulla and contributes to inhibition of voluntary movements, and also reduces muscle tone.
Vestibulospinal tracts
The vestibulospinal tracts do not decussate.
There are two vestibulospinal tracts that control anti-gravity muscles via LMNs:
- Medial vestibulospinal tract: originates in the medial vestibular nucleus, to control ipsilateral postural and tone adjustments in response to the vestibular apparatus.
- Lateral vestibulospinal tract: originates in the lateral, superior and inferior vestibular nuclei, to control ipsilateral postural and tone adjustments in response to the vestibular apparatus.
Rubrospinal tract
The rubrospinal tract decussates.
The rubrospinal tract begins in the red nucleus, where fibres immediately decussate and descend through the pons and medulla and into the spinal cord.
It is thought that the rubrospinal tracts supply upper limb flexors as well as trunk flexors. Disinhibition of the rubrospinal tract leads to upper limb flexion.
Inhibition of the rubrospinal tract leads to upper limb extension.
Tectospinal tract
The tectospinal tract decussates.
The tectospinal tract begins in the tectum, or roof of the midbrain, where the superior and inferior colliculi are located. Collectively, the two superior and two inferior colliculi are referred to as corpora quadrigemina.
The superior colliculus is involved in reflexive responses to visual stimuli.
The inferior colliculus is involved in reflexive responses to auditory stimuli.
Together, the colliculi send information about sights and sounds to the tectospinal tract, which decussates soon after leaving these structures, to supply muscles of the head and neck for reflexive localisation of these stimuli.
Clinical relevance: UMN and LMN lesions of the body
Pathologies of both UMN and LMN present with different clinical signs that we can detect on clinical examination.
Table 3. UMN vs LMN lesions
| Physical sign | UMN lesion | LMN lesion |
| Tone | Hypertonia | Hypotonia |
| Paralysis (or paresis) | Spastic | Flaccid |
| Fasciculations | Absent | Present |
| Atrophy | Minimal | Marked |
| Reflexes | Exaggerated/clonus | Diminished/absent |
| Babinski’s sign | Present | Absent |
You will notice that UMN lesions present with hypertonia and spastic paralysis, whereas LMN lesions are usually associated with hypotonia and flaccid paralysis. This is because of the impaired ability for motor neurons to regulate descending signals, giving rise to disordered spinal reflexes.
To understand more, we must integrate what we know about the central nervous system: as much as it is involved with activating pathways, it can also suppress pathway activity. That is, the corticospinal tract also helps in conscious inhibition of muscle (lack of the signal). If we sever the UMNs of the corticospinal tract, there is a loss of inhibitory tone of muscles. The effect of this is two-fold (on a simple level; there is much more to it!):
- No LMN inhibition leads to LMN activation and constant contraction of muscles; and
- Incoming sensory afferents carrying muscle stretch information are not received, and our body thinks we are not compensating and signals are amplified.
This leads to the hypertonia and spastic paralysis we see in UMNs. If LMNs are damaged or lost, there is nothing to tell the muscles to contract, with resultant hypotonia and flaccid paralysis.
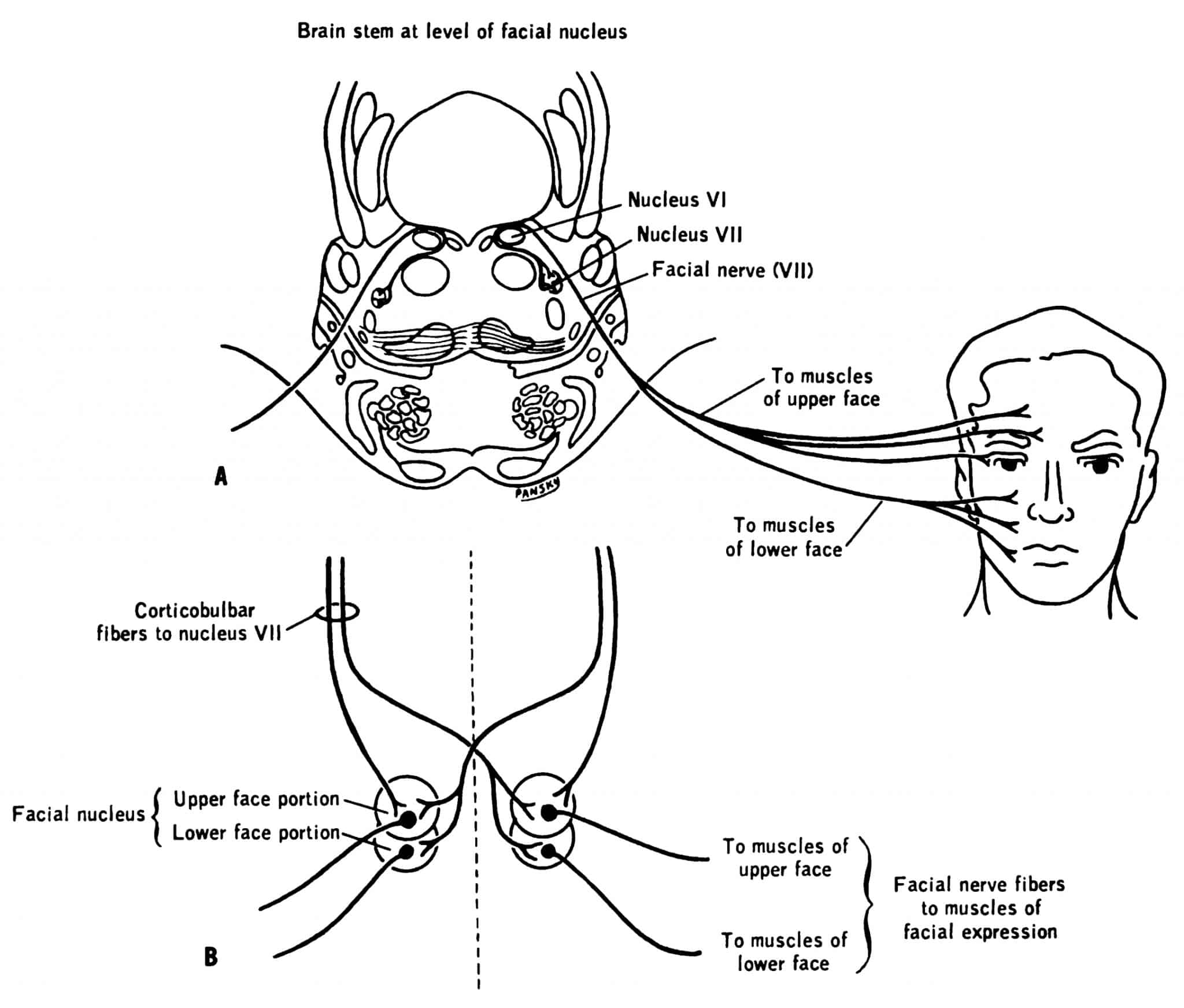
Clinical relevance: UMN and LMN lesions of the facial nerve
The facial nerve presents a classic example of UMN versus LMN lesions. We have discussed that the upper half of the face receives a bilateral cortical supply, whereas the lower half of the face receives contralateral cortical supply only.
Knowing this, we can tell whether someone has an UMN or LMN lesion to the facial nerve.
We can also tie in the signs seen in UMN versus LMN lesions. The paralysis in an UMN facial lesion will classically be spastic. The paralysis of a LMN facial lesion is flaccid.
Table 4. UMN vs LMN facial nerve lesions
| Facial field | UMN | LMN |
| Upper half | Preserved | Paresis or paralysis |
| Lower half | Paresis or paralysis | Paresis or paralysis |
Clinical relevance: decerebrate and decorticate posturing
Now that we understand the rubrospinal tract and the role it plays in adjusting flexor tone in the upper limb, we can discuss decorticate versus decerebrate posturing. Both types of posture involve lower limb extension.
Decerebrate posturing refers to an adopted posture of upper limb extension. This occurs when a lesion below the red nucleus prevents the red nucleus from activating the upper limb flexors, resulting in upper limb extension.
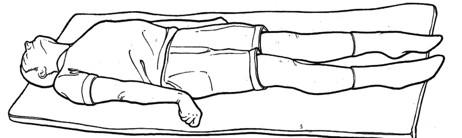
Decorticate posturing refers to an adopted position of upper limb flexion. This occurs when a lesion above the red nucleus prevents inhibitory tone of the red nucleus, allowing it to cause flexion of the upper limb.
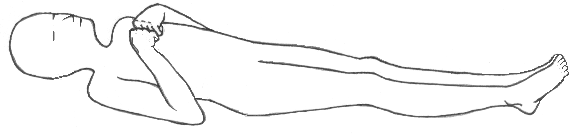
The other extrapyramidal tracts do play a role in this:
- The rubrospinal and lateral reticulospinal tracts together have a bias towards flexion, and the input from these two tracts outweighs the postural control of the medial reticulospinal and medial/lateral vestibulospinal tracts in the upper limbs.
- This results in upper limb flexion in decorticate posturing for lesions above the red nucleus.
- In decerebrate posturing, the loss of the rubrospinal tract causes the lateral reticulospinal tract to be overwhelmed by the other extrapyramidal tracts, resulting in upper limb extension.
- Disruption of the lateral corticospinal tract allows the medial reticulospinal and medial and lateral vestibulospinal tracts of the lower limbs to overcome the input from the lateral reticulospinal tract.
- This results in lower limb extension in both decorticate or decerebrate posturing.
References
Reference texts
- Sinnatamby, C. S. (2011). Last’s Anatomy, International Edition: Regional and Applied. Elsevier Health Sciences.
- Moore, K. L., Dalley, A. F., & Agur, A. M. (2013). Clinically oriented anatomy. Lippincott Williams & Wilkins.
- Nolte, J. (2002). The human brain: an introduction to its functional anatomy.
- Snell, R. S. (2010). Clinical neuroanatomy. Lippincott Williams & Wilkins.
Reference images
- OpenStax College. License: [CC BY]
- BruceBlaus. License: [CC BY-SA]
- Polarlys and Mikael Häggström. License: [CC BY-SA]
- CueFlash. License: [Public domain]
- Delldot at English Wikipedia. License: [Public domain]
- House, Earl Lawrence. Pansky, Ben. License: [Public domain]
